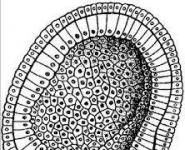
Строение ресничек и жгутиков клетки. Реснички и жгутики: сжатая характеристика, строение и роль в клетках Жгутики у бактерий являются органами
Строение. Около половины известных видов бактерий на поверхности имеют органы движения - волнообразно изогнутые жгутики. Масса жгутиков составляет до 2 % сухой массы бактерии. Длина жгутика больше длины тела микроорганизма и составляет 3–12 мкм; толщина жгутика 0,02 мкм, причем полярные жгутики более толстые, чем перитрихиальные.
Жгутики состоят из белка флагеллина (лат. flagella - жгутик), который по своей структуре относится к сократительным белкам типа миозина. В составе жгутика имеется либо одна гомогенная белковая нить, либо 2–3 нити, плотно свернутые в косу. Нить жгутика - жесткая спираль, закрученная против часовой стрелки; шаг спирали специфичен для каждого вида бактерий.
Число, размеры и расположение жгутиков являются признаками, постоянными для определенного вида, и учитываются при систематике. Однако у некоторых бактерий могут образовываться жгутики разных типов. Кроме того, наличие жгутиков зависит от условий внешней среды: на твердых средах при длительном культивировании бактерии могут утратить жгутики, а на жидких - вновь приобрести. Количество и расположение жгутиков у одного и того же вида может определяться стадией жизненного цикла. Следовательно, не стоит переоценивать таксономическое значение этого признака.
Классификация бактерий по числу и расположению жгутиков:
1. Атрихи - жгутики отсутствуют.
2. Монотрихи - один жгутик, расположенный на одном из полюсов клетки (род Vibrio) - монополярное монотрихальное расположение жгутиков, самые подвижные бактерии.
3. Политрихи - много жгутиков:
– лофотрихи - пучок жгутиков на одном полюсе клетки(роды Pseudomonas , Burkholderia )-монополярное политрихальное расположение жгутиков;
– амфитрихи - на каждом полюсе клетки расположено по пучку жгутику (род Spirillum) - биполярное политрихальное расположение жгутиков;
– перитрихи - жгутики расположены без определенного порядка по всей поверхности клетки (сем. Enterobacteriaceae (роды Escherichia, Proteus ), сем. Bacillaceae, сем. Clostidiaceae ), число жгутиков от 6 до 1000 на клетку в зависимости от вида бактерий (рис. 7).
Рис.7. Варианты расположения жгутиков у бактерий:
1 - монотрих, 2 - лофотрих;
3 - амфитрих; 4 - перитрих.
– При электронной микроскопии обнаружено, что жгутик состоит из трех частей: спиральной нити, крюка и базального тела (рис. 8).
Основную часть жгутика составляет длинная спиральная нить (фибрилла) - жесткий полый цилиндр диаметром около 120 нм, состоящий из белка флагеллина. По длине нити белковые молекулы образуют 11 рядов и уложены в виде спирали. В процессе роста нити белковые молекулы, синтезированные внутри клетки, проходят через полость цилиндра и пристраиваются в спираль на ее конце. На конце жгутика имеется белковая шапочка (крышечка), закрывающая отверстие цилиндра и препятствующая выходу молекул белка в окружающую среду. Длина нити жгутика может достигать нескольких микрометров. У некоторых видов бактерий жгутик снаружи дополнительно покрыт чехлом. У поверхности КС спиральная нить переходит в утолщенную изогнутую структуру - крюк.
Рис. 8. Схема строения жгутика
2. Крюк (толщиной 20–45 нм) вблизи поверхности клетки - относительно короткий цилиндр, состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом.
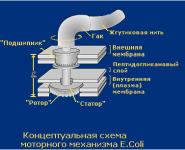
3. Базальное тело находится в основании жгутика и обеспечивает его вращение. Базальное тело содержит 9–12 различных белков и состоит из двух или четырех дисков (колец), нанизанных на стержень, являющийся продолжением крюка. Эти кольца вмонтированы в ЦПМ и КС. Два внутренних кольца (M и S) - обязательные составные части базального тела. M-кольцо локализовано в ЦПМ, S-кольцо располагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных бактерий. Два наружных кольца (D и L) необязательны для движения, так как имеются только у грамотрицательных бактерий, локализованы соответственно в пептидогликановом слое и в наружной мембране КС. Кольца S, D и L неподвижны и служат для фиксации жгутика в КС. Вращение жгутика определяется вращением M–кольца, встроенного в ЦПМ клетки. Таким образом, особенности строения базального тела жгутика определяются строением КС.
Функционально базальное тело представляет собой электромотор, работающий на протонах. М–кольцо базального тела (вращающийся ротор), окружено мембранными белками, имеющими отрицательные заряды (статор мотора). Бактериальная клетка обладает эффективным механизмом, позволяющим превращать электрохимическую энергию в механическую. Поэтому на работу жгутика бактерия тратит около 0,1 % всей расходуемой ею энергии. При работе жгутика используется протондвижущая сила, которая обеспечивается разностью концентраций протонов на внешней и внутренней сторонах мембраны (на внешней стороне их больше) и наличием более отрицательного заряда на внутренней стороне мембраны. Протондвижущая сила заставляет протоны проходить через базальное тело внутрь клетки, при этом они задерживаются на определенных участках ротора, придавая им положительный заряд, затем протоны уходят внутрь клетки. Заряженные участки расположены таким образом, что возникает сила притяжения между заряженными участками ротора и статора, М-кольцо начинает вращаться со скоростью около 300 об/c. Механизм вращения: зарядка–перезарядка группы COOH в аминокислотах. Для полного оборота кольца через базальное тело должно пройти 500–1000 протонов. Вращение М-кольца через жестко связанную с ним ось и крюк передается нити жгутика, которая функционирует как пропеллер или корабельный винт. Бактерия плывет до тех пор, пока работает винт, вклад инерции исключительно мал.
Кроме того, бактерии, даже мертвые, находящиеся в водной среде, перемещаются в результате броуновского движения. Бактериальная клетка все время подвержена ударам окружающих молекул, находящихся в тепловом движении. Удары, наносимые с разных сторон, бросают бактерию из стороны в сторону.
Тип движения жгутиков - вращательный. Существуют два вида движения: прямолинейное и кувыркание (периодические случайные изменения направления движения). Когда жгутики вращаются против хода часовой стрелки (около 1 секунды), с частотой 40–60 об./с (близко к скорости среднего электромотора), их нити сплетаются в единый жгут (рис. 9а). Вращение жгутиков передается клетке. Так как клетка намного массивнее жгутика, она начинает двигаться по прямой в противоположном направлении, со скоростью в 3 раза меньшей, чем скорость движения жгутика.
Так обеспечивается поступательное движение клетки, скорость которого в жидкой среде для разных видов бактерий составляет 20–200 мкм/с (это соответствует примерно 300–3000 длин тела в минуту) и более медленное перемещение по поверхности твердых сред.
Плыть целенаправленно в одном направлении бактерия может не более 3 с, затем удары окружающих молекул разворачивают ее в случайном направлении. Поэтому бактерии выработали механизм спонтанного изменения направления движения - переключение жгутикового мотора. Когда он начинает вращаться по ходу часовой стрелки (около 0,1 с), бактерия останавливается и переворачивается (совершает «кувырок») в случайном направлении. При этом жгутики разлетаются в разные стороны (рис. 9б). У амфитрихов при движении один пучок жгутиков вывернут наизнанку (подобно вывернутому ветром зонту). Затем мотор снова переходит к вращению против часовой стрелки, и бактерия опять плывет по прямой, но уже в другом, случайном направлении.
Жгутики могут также изменять направление движения в ответ на внешний стимул. Если бактерия перемещается в сторону оптимальной концентрации аттрактанта, жгутики проталкивают клетку через среду, ее прямолинейное движение становится более длительным, а частота кувырканий более низкой, что позволяет ей в конечном итоге перемещаться в нужном направлении.
Известны случаи существования бездействующих (парализованных) жгутиков. Для движения жгутиковых бактерий необходима интактность (неповрежденность) КС. Обработка клеток лизоцимом, приводящая к удалению пептидогликанового слоя КС, вызывает потерю способности бактерий к движению, хотя жгутики остаются при этом неповрежденными.
Таксисы бактерий. Пока окружающая среда остается неизменной, бактерии плавают беспорядочно. Однако совершенно однородной окружающая среда бывает редко. Если среда неоднородна, бактерии проявляют элементарные поведенческие реакции: активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие генетически детерминированные целенаправленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай - аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис.
Хемотаксис - движение в определенном направлении относительно источника химического вещества. Химические вещества делят на две группы: инертные и вызывающие таксисы - хемоэффекторы. Среди хемоэффекторов есть вещества, привлекающие бактерий, - аттрактанты (сахара, аминокислоты, витамины, нуклеотиды), и вещества, их отпугивающие, - репелленты (некоторые аминокислоты, спирты, фенолы, неорганические ионы). Аттрактантом для аэробных и репеллентом для анаэробных прокариот является молекулярный кислород. Аттрактанты часто представлены пищевыми субстратами, хотя не все вещества, необходимые для организма, выступают в качестве аттрактантов. Также не все ядовитые вещества служат репеллентами и не все репелленты вредны. Таким образом, бактерии способны реагировать не на любые соединения, а только на определенные и различные для разных бактерий.
В поверхностных структурах бактериальной клетки есть специальные белковые молекулы - рецепторы, специфически соединяющиеся с определенным хемоэффектором, при этом молекула хемоэффектора не изменяется, а в молекуле рецептора происходят конформационные изменения. Рецепторы расположены неравномерно по всей поверхности клетки, а сконцентрированы на одном из полюсов. Состояние рецептора отражает внеклеточную концентрацию соответствующего эффектора.
Хемотаксис имеет приспособительное значение. Например, формы холерного вибриона с нарушенным хемотаксисом оказываются менее вирулентными.
Аэротаксис - нуждающиеся в молекулярном кислороде бактерии скапливаются вокруг пузырьков воздуха, попавших под покровное стекло.
Фототаксис - движение к свету или от него, свойствен фототрофным бактериям, использующим свет в качестве источника энергии.
Магнитотаксис - способность водных бактерий, содержащих кристаллики железосодержащих минералов, плыть вдоль линий магнитного поля Земли.
Термотаксис - движение в сторону изменения температуры, что имеет большое значение для некоторых патогенных бактерий.
Вискозитаксис - способность реагировать на изменение вязкости раствора. Обычно бактерии стремятся в среду с большей вязкостью, что имеет большое значение для патогенных видов.
Скольжение бактерий. Способность к скольжению с небольшой скоростью (2–11 мкм/с) по твердому или вязкому субстрату обнаружена у некоторых прокариот, напр., микоплазм.
Существуют несколько гипотез, объясняющих скользящее движение. Согласно гипотезе реактивного движения оно обусловлено выделением слизи через многочисленные слизевые поры в КС, в результате чего клетка отталкивается от субстрата в направлении, противоположном направлению выделения слизи. Согласно гипотезе «бегущей волны» скользящее движение у подвижных безжгутиковых форм связано с наличием между пептидогликановым слоем и наружной мембраной КС тонкого белкового слоя из упорядоченно расположенных фибрилл, аналогичных нитям жгутиков. Вращательное движение фибрилл, «запускаемое» этими структурами, приводит к появлению на поверхности клетки «бегущей волны» (движущихся микроскопических выпуклостей КС), в результате чего клетка отталкивается от субстрата. Наконец, у некоторых скользящих бактерий описаны структуры, напоминающие базальные тела жгутиковых форм.
Функции жгутиков:
1. Обеспечивают адгезию - начальную стадию инфекционного процесса.
2. Обеспечивают подвижность бактерий.
3. Определяют антигенную специфичность, это Н-антиген.
Выявление жгутиков:
1. Фазовоконтрастная микроскопия нативных препаратов («раздавленной» и «висячей» капли). Микроскопически подвижность определяют у клеток суточной культуры. Для того чтобы отличить подвижность от пассивного броуновского движения, к капле исследуемой культуры добавляют каплю 5 %–ного водного раствора фенола, активное движение в этом случае прекращается.
2. Темнопольная микроскопия нативных препаратов.
3. Световая микроскопия окрашенных красителями или металлами препаратов. Так как жгутики очень легко повреждаются при приготовлении препарата, в повседневной практике эти методы используется редко.
Для окраски жгутиков используют клетки, выращенные на скошенном агаре. Бактериальной петлей отбирают клетки, находящиеся у конденсационной воды и осторожно переносят в стерильную дистиллированную воду такой же температуры, что и температура инкубирования бактерий на скошенном агаре, а бактерии с петли не стряхивают, а осторожно погружают в воду. Пробирку с бактериями оставляют при комнатной температуре на 30 мин. Используют химически чистое (вымытое в хромовой смеси) стекло, на которое наносят 2–3 капли суспензии. Суспензию распределяют по поверхности стекла, осторожно его наклоняя. Высушивают препарат на воздухе.
Жгутики очень тонкие, поэтому их можно обнаружить только при специальной обработке. Вначале при помощи протравки достигается разбухание и увеличение их размера, а затем производится окраска препарата, благодаря чему они становятся видимыми при световой микроскопии.
Чаще используют метод серебрения по Морозову (рис. 10):
– препарат фиксируют раствором ледяной уксусной кислоты 1 минуту, промывают водой;
– наносят раствор таннина (дубящий, делающий жгутики более плотными) на 1 мин, промывают водой;
– обрабатывают препарат при подогревании импрегнирующим раствором азотнокислого серебра 1–2 мин, промывают водой, высушивают и микроскопируют.
При микроскопии видны темно-коричневые клетки и более светлые жгутики.
Рис. 10. Выявление жгутиков методом серебрения
Рис. 11. Выявление жгутиков
методом электронной микроскопии
4. Электронная микроскопия препаратов, напыленных тяжелыми металлами (рис. 11).
5. Косвенно - по характеру роста бактерий при посеве в полужидкий 0,3 %–ный агар. После инкубирования посевов в термостате в течение 1–2 сут отмечают характер роста бактерий:
– у неподвижных бактерий (напр., S. saprophyticus ) наблюдается рост по ходу укола - «гвоздь», а среда прозрачна;
– у подвижных бактерий (напр., Е. со1i )наблюдается рост в стороны от укола, по всему столбику агара - «елочка», и диффузное помутнение среды.
Все бактерии подразделяются на подвижные и неподвижные. Органами движения у бактерий являются жгутики. Они состоят из белка флагеллина, который по своей структуре относится к сократительным белками типа миозина.
Основанием жгутика
является базальное тельце, состоящее из системы дисков (блефаропласта: 1 диск - наружная сторона клеточной стенки, 2 диск - внутренняя сторона клеточной стенки, 3 диск - цитоплазматическая мембрана), "вмонтированных" в цитоплазматическую мембрану и клеточную стенку. Длина жгутика больше длины тела самого микроба.
По числу жгутиков
и их расположению подвижные микроорганизмы подразделяются на:
1. Монотрихи, имеющие на конце тела один жгутик (самые подвижные). Например, Vibrio cholerae.
2. Лофотрихи, имеющие пучок жгутиков на одном из полюсов клетки. Например, Burkholderia (Pseudomonas) pseudomalei - возбудитель мелиоидоза.
3. Амфитрихи, имеющие жгутик на обоих полюсах клетки. Например, Spirillum volutans.
4. Перитрихи, имеющие жгутики по всему периметру клетки. Например, Escherichia coli, Salmonella typhi.
Выявление жгутиков. Жгутики очень тонкие, поэтому их можно обнаружить только при специальной обработке. В частности, вначале при помощи протравы достигается разбухание и увеличение их размера, а затем производится окраска препарата, благодаря чему они становятся видимыми при световой микроскопии. Выявлять жгутики можно окраской по Морозову, Леффлеру, а также электронной микроскопией. Обнаружить жгутики можно и по активной подвижности бактерий.
Движение микробов наблюдают в препаратах "раздавленной" и "висячей" капли из живых культур. Микроскопируют эти препараты сухим или иммерсионным объективом в темном поле или в фазовом контрасте. Кроме того, подвижность можно определить по характеру роста бактерий в полужидком агаре.
Пили у бактерий.
Пили (pili), синонимы: ворсинки, фимбрии, - тонкие полые нити белковой природы, покрывающие поверхность бактериальных клеток. В отличие от жгутиков не выполняют двигательную функцию.
Пили отходят от поверхности клетки и состоят из белка пилина .
По своему функциональному назначению подразделяются на 2 типа.
1) Пили первого типа имеются у большинства бактерий, поэтому они получили название "ворсинки общего типа" (common pili). Обусловливают прикрепление или адгезию бактерий к определенным клеткам организма хозяина. Адгезия является первоначальной стадией любого инфекционного процесса.
2) Пили второго типа (синонимы: конъюгативные, или половые - sex pili) имеются только у бактерий-доноров, имеющих специальную плазмиду. Их количество невелико - 1-4 на клетку.
Половые пили выполняют следующие функции:
1. Участвуют в передаче генетического материала от одной клетки к другой при конъюгации бактерий.
2. На них адсорбируются специфические вирусы бактерий – бактериофаги
Споры бактерий, условия образования, расположение, механизм и этапы окраски по Ауески.
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки.
Спорообразование - это способ сохранения вида (генофора) во внешней среде при неблагоприятных условиях, а не способ размножения.
Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). Внутри бактериальной клетки образуется одна спора (эндоспора).
Стадии спорообразования
1. Подготовительная. В цитоплазме бактерий образуется уплотненный участок, не имеющий свободной воды, называемый "спорогенной зоной", в которой содержится нуклеоид.
2. Стадия предспоры (проспоры). Вокруг спорогенной зоны образуется оболочка из двойной цитоплазматической мембраны.
3. Образование кортекса, состоящего из пептидогликана и наружной мембраны с повышенным содержанием солей кальция и липидов.
4. Стадия созревания. С внешней стороны наружной мембраны образуется оболочка споры, после чего вегетативная часть клетки лизируется, освобождая спору.
Для передвижения в водной среде некоторые микроорганизмы используют жгутиковидный орган – «жгутик». Этот орган, встроенный в мембрану клетки, позволяет микроорганизму по желанию передвигаться в выбранном им направлении с определённой скоростью.
Мужские половые клетки также используют жгутик для передвижения.
Определенной время учёные знали о жгутиках. Однако, знания об их структурных особенностях, которые появились лишь за последнее десятилетие или около того, явились для них огромным сюрпризом. Было установлено, что жгутик двигается посредством очень сложного «органического двигателя», а не простого вибрирующего механизма, как считалось раньше.
Этот двигатель сформирован по тем же механическим принципам, что и электрический двигатель. В нём есть две главные части: движущаяся часть («ротор») и станционарная часть («статор»).
Бактериальный жгутик отличается от всех органических систем, которые совершают механические движения. Клетка не использует имеющуюся в наличии энергию, сохраняемую в молекулах АТФ. Вместо этого, она имеет специальный энергетический ресурс: микроорганизм использует энергию потока ионов сквозь их внешние мембраны. Внутренняя структура двигателя чрезвычайно сложна. В создании жгутика принимают участие около 240 различных белков. Каждый из них занимает определённое место. Учёные установили, что эти белки проводят сигналы, включающие и выключающие двигатель, формируют соединения, облегчающие движения на атомном уровне, и активизируют другие белки, которые присоединяют жгутик к клеточной мембраны. Моделей, разработанных для резюмирования работы системы, вполне достаточно для описания сложной структуры системы. (1)

Сложной структуры бактериального жгутика самой по себе уже достаточно, чтобы опровергнуть теорию эволюции, поскольку жгутик имеет неснижаемо сложную структуру. Даже если одна единственная молекула этой невероятно сложной структуры исчезнет или повредится, жгутик не будет ни работать, ни представлять пользу для микроорганизма. Жгутик должен идеально работать с самого первого момента его существования. Этот факт ещё раз подчёркивает абсурдность утверждения теории эволюции о «ступенчатом развитии».

Даже у тех созданий, которых эволюционисты считают «простейшими», существует потрясающая структура. Бактериальный жгутик является одним из бесчисленных примеров. Этот микроорганизм передвигается в воде, двигая этим органом на своей оболочке. Когда были изучены внутренняя система этого хорошоизвестного органа, учёные всего мира были удивлены, обнаружив, что микроорганизм имеет чрезвычайно сложный по своему строению электрический двигатель. Этот электрический двигатель, который включает около пятидесяти разнообразных молекулярных субедениц, имеет достаточно замысловатую структуру, что изображено ниже.
Бактериальный жгутик является очевидным доказательством того, что даже предположительно «примитивные» создания имеют необычное строение. По мере того, как человечество всё более постигает детали, становится очевидным, что те организмы, которые учёные 19-го столетия, включая Дарвина, считали простейшими, на самом деле так же сложны, как и другие. Другими словами, с приходом понимания о совершенстве создания, очевидным становится бессмысленность попыток найти сотворению альтернативное объяснение.

Микороорганизм плавает в вязкой жидкой среде, вращая спиралевидными пропеллерами, называемыми жгутиками .
Бактериальный жгутик является наномашиной, состоящей из 25 разнообразных белков, в количестве от нескольких штук до десятков тысяч. Он состоит из собрания этого большого количества белков, каждый из которых в разных частях выполняет определённую функцию, такую как ротация двигателя, изоляция, приводной вал, очерёдность переключения регулятора, универсальная связка, спиралевидный пропеллер, и ротативный усилитель для самосборки.
Жгутиковые протеины синтезируются внутри тела клетки и транспортируются по длинному, узкому центральному каналу в жгутике к его периферическому (внешнему) окончанию, где они с помощью жгутикового наконечника в качестве установочного двигателя, эффективно и самостоятельно могут создавать сложные структуры наноразмером. Вращательный двигатель, диаметр которого всего от 30 до 40 нм, производит вращение жгутика с частотой около 300 Гц и мощностью в 10-16 Вт, с эффективностью преобразования энергии близкой к 100%.
Структурные дизайны и функциональные механизмы, обнаруженные в сложном механизме бактериального жгутика, могут обеспечить человечество множеством новаторских технологий, которые станут основой для будущей нанотехнологии, для которой мы сможем найти много полезных способов применения.(2)
Реснички и жгутики содержат высокоупорядоченную структуру, называемую аксонемой
Аксонема состоит из девяти наружных дуплетов микротрубочек, окружающих пару центральных микротрубочек
Радиальные спицы представляют собой комплекс из нескольких полипептидов, которые связывают каждый наружный дуплет с центром аксонемы
С каждым внешним дуплетом связываются динеины, домены моторов которых распространяются на примыкающие внешние дуплеты
Динеин сдвигает внешние дуплеты по отношению друг к другу; структурные связи между другими дуплетами переводят скольжение в наклон аксонемы
Кинезины участвуют в сборке жгутиков, транспортируя белки аксонемы в отдаленный конец жгутика
Неподвижные первичные реснички участвуют в сенсорных процессах
Наряду с транспортировкой карго в клетках, микротрубочки также участвуют в перемещении клеток в окружающей среде. Это перемещение осуществляется с помощью ресничек и жгутиков , длинных и тонких выростов, которые отходят как волоски на поверхности многих клеток. Каждая такая органелла состоит из длинного пучка микротрубочек, окруженного выростом плазматической мембраны. Взаимодействие между микротрубочками вызывает изгиб структуры, который возможен в разных направлениях. В результате этих движений жидкость может продвигаться вдоль поверхности клеток, как показано на рисунке ниже. Для группы неподвижных клеток, например эпителиальных, это позволяет жидкой среде и различным объектам перемещаться вдоль поверхности ткани. В случае изолированных, неприкрепленных клеток они сами движутся в жидкости (т. е. плавают). Реснички и жгутики присутствуют у многих одноклеточных организмов, таких как Paramecium и Chlamydomonas (зеленая водоросль), а также находятся на сперматозоидах у большинства эукариот. У млекопитающих реснички покрывают апикальные участки некоторых эпителиальных клеток и совершают синхронные движения. При этом возникают волны движения ресничек, которые проходят по поверхности ткани. В трахее это движение используется для удаления слизи и дебриса из дыхательных путей; в яйцеводе оно обеспечивает транспортировку яйцеклетки из яичника в матку, а в мозгу создает циркуляцию цереброспинальной жидкости.
Реснички и жгутики обладают одинаковой структурой и механизмом подвижности, однако в некоторых отношениях они различаются. Наиболее существенные различия касаются длины, их количества в клетке и характера движения. Реснички короче (10-15 мкм) и количество их на клетку достигает 100 или больше. Каждая ресничка генерирует усилие за счет наклона у основания. Внешняя часть реснички остается жесткой и перегиб у основания сдвигает ее в движение, напоминающее гребок весла в воде. После этого наступает восстановительный размах, при котором перегиб реснички распространяется от основания на конец, подготавливая ресничку к следующему циклу движения. На рисунке ниже представлено движение реснички Для того чтобы проследить за отдельными стадиями движения, для регистрации использовали замедленную видеосъемку. Движения реснички столь интенсивны (много наклонов в секунду), что при съемке в обычном режиме она выглядит нечетко.
Обычно длиннее (10-200 мкм), чем реснички, и обычно клетка содержит только один или два жгутика. Они также генерируют силу при изгибе; при этом, как показано на рисунке ниже, от основания к вершине жгутика распространяется S-образная волна. Для ресничек и жгутиков характерен общий механизм движения, в основе которого лежит изгибание структуры. Различные пути распространения изгиба вдоль органеллы обусловливают различные формы волн, генерируемых ресничками и жгутиками. Поскольку два типа органелл представляют собой вариации на одну и ту же тему, мы рассмотрим их общие свойства и будем пользоваться термином жгутик для описания структуры и подвижности обеих органелл, если специально не оговаривается, что речь идет о ресничках.
При отделении от клетки жгутики продолжают биение. Это говорит о том, что оно обеспечивается самой органеллой. После удаления плазматической мембраны изолированные жгутики также продолжают биться при условии, что у них сохранился запас АТФ. Эти данные свидетельствуют о том, что движения обеспечиваются белковым содержимым жгутика и энергией гидролиза АТФ.
Внутренний белковый компонент жгутика представляет собой упорядоченную структуру, состоящую по крайней мере из 250 различных полипептидов. Эта структура называется аксонемой. Структура аксонемы жгутиков одинакова для столь различных организмов, как одноклеточное простейшее Chlamydomonas и человек.
Одноклеточная водоросль Chlamydomonas reinhardtii в световом и флуоресцентном микроскопах.В верхней части клетки видны два больших жгутика.
Во флуоресцентном микроскопе микротрубочки флуоресцируют красным; показывая, что жгутики состоят из .
Клетки Chlamydomonas способны передвигаться в воде за счет биений жгутиков.
Строение аксонемы представлено на рисунке ниже. Наиболее примечательным элементом структуры, особенно заметным на поперечном срезе, являются определенным образом организованные дуплеты микротрубочки, расположенные вдоль всей аксонемы. Девять необычных «двойных микротрубочек» расположены по кругу. Каждая пара состоит из одной обычной микротрубочки, содержащей 13 протофиламентов (т. н. А трубочка) и одной неполной микротрубочки (В трубочка). Эта микротрубочка состоит из 10-11 протофиламентов и связана со стенкой А трубочки. В центре кольца, образованного дуплетами микротрубочек, находятся две обычных микротрубочки, содержащие 13 протофиламентов («центральная пара»). Такое расположение микротрубочек в аксонеме сокращенно описывается как «9+2». Все микротрубочки обладают одинаковой полярностью, их плюс-концы направлены к кончику жгутика, а минус-концы - к его основанию. С микротрубочками связываются различные белки, оказывающие на них стабилизирующее действие.
В аксонеме микротрубочки связаны между собой множеством различного типа связей. Белки, которые формируют эти связи, необходимы для организации микротрубочек в единую согласованную систему, способную двигаться и координировать эти движения для придания им волнового характера. Соседние дуплеты микротрубочек соединены между собой по периферии аксонемы белком, называемым нексином. Дуплеты микротрубочек также связаны с микротрубочками, расположенными в центре, с помощью полипептидных комплексов, которые образуют как бы спицы с видимыми головками. Эти структуры имеют сложное строение: спицы и их головки содержат 17 различных полипептидов. Головки расположены вокруг внутренней оболочки, структуры, окружающей две центральные микротрубочки. Усилие в аксонеме генерируется с помощью аксонемных динеинов (их также называют «ресничатыми» или «жгутиковыми» динеинами). Эти белки связывают между собой соседние дуплеты микротрубочек; хвостовой домен связывается с А трубочкой в одного дуплета, а головной домен с В трубочкой следующего дуплета. Различные связи, образованные нексином, спицами и динеинами, расположены через правильные промежутки по всей длине аксонемы, однако они характеризуются разной периодичностью. Поэтому все три компонента трудно наблюдать в электронном микроскопе на препаратах, представляющих собой поперечные срезы аксонем. Однако, когда удается видеть все три структурных элемента, они напоминают колесо с массивными спицами и втулкой.
Структура и расположение динеинов в остальной части аксонемы представляются сложными. В аксонеме содержится более одной формы динеина, каждая из них достаточно велика и состоит из большего числа различных полипептидов по сравнению с динеином, содержащимся в цитоплазме. Различные формы содержат один, два иди три моторных домена, и расположены в разных местах аксонемы. Смежные дуплеты микротрубочки связаны двумя наборами молекул динеина, которые называются внутренними и внешними ручками. Внешние ручки содержат только динеины с двумя или тремя головками, а белки с одной или двумя головками находятся во внутренних ручках.
Каким образом все эти связи обеспечивают подвижность жгутико в и задают соответствующий характер биения? Основной вопрос заключается в том, как функционируют динеины в этой структуре, поскольку они представляют собой моторы и с них должно начинаться движение. Для того чтобы выяснить роль динеина в обеспечении подвижности, необходимо отделить жгутики от клеток и удалить мембрану аксонемы. После этого аксонемы подвергают кратковременной обработке протеазой для того, чтобы разрушить нексиновые связи между внешними дуплетами микротрубочек. Если затем к препарату аксонем добавить АТФ, то микротрубочки сдвигаются по отношению к друг другу. Этот эффект вызывается динеинами, хвосты которых связаны с дублетом микротрубочек, генерирующих усилие в направлении от плюс- к минус-концу соседней пары. В интактной аксонеме динеин не может сдвигать внешние дублеты, поскольку они связаны между собой с помощью нексина. Поэтому усилие, генерируемое динеином, приводит к сгибанию жгутика.
Реснички и жгутики генерируют биение за счет распространения сгибательного движения по аксонеме. Сгибание начинается в основании реснички или жгутика и распространяется по направлению к дистальному концу. Сгибание происходит потому, что динеин активен только в пределах небольшого участка аксо-немы. Для того чтобы обеспечить сгибание, динеины активируются последовательно как вдоль длины, так и по периферии аксонемы. Активность динеинов контролируется центральными микротрубочками и спицами; жгутики мутантных клеток, у которых эти структуры отсутствуют, не способны совершать биение. У некоторых организмов центральные микротрубочки быстро вращаются, и в это время они могут передавать сигналы на спицы, которые, в свою очередь, активируют динеин. В центральных микротрубочках и в спицах находится несколько киназ и фосфатаз. Предполагается, что при вращении центральных трубочек запускается локальная система передачи сигнала, которая активирует динеины. Посредством быстрой локальной активации и инактивации специфических изоформ динеина аксонемы генерируют движения и регулируют силу и частоту биений.
В основании жгутика находится структура, которая называется базальным тельцем. Эти структуры обладают таким же строением, как и центриоли. Каждое базальное тело представляет собой цилиндр, состоящий из 9 триплетов микротрубочек, каждая из которых состоит или из 13 (трубочка А) или из 11 протофиламентов (трубочки В и С). Трубочки А и В служат матрицами при сборке 9 наружных дуплетов микротрубочек аксонемы. Базальное тело остается связанным с основанием аксонемы и служит для прикрепления последней к телу клетки.
Как происходит сборка жгутика , можно выяснить, удалив его с поверхности клетки и проследив образование нового. Жгутик регенерирует менее чем за час и в ходе этого процесса функционирует (т. е. осуществляет биения). Рост нового жгутика происходит на плюс-концах микротрубочек аксонемы, расположенных в дистальных участках каждого жгутика. Сборка жгутика предполагает, что необходимые компоненты аксонемы должны транспортироваться на концы структур и по мере роста собираться в аксонемы. Транспорт осуществляется в больших белковых комплексах, которые движутся в направлении кончика жгутика по наружной поверхности аксонемы, расположенной непосредственно под плазматической мембраной. Это движение называется интрафлагеллярным транспортом (IFT) и осуществляется с участием кинезина. Белковые комплексы также могут двигаться от вершины жгутика к его основанию (к минус-концам микротрубочек), однако функциональное значение этого направления транспорта неизвестно. IFT по направлению к телу клетки происходит за счет цитоплазматического динеина.
Хотя большинство ресничек представляют собой подвижные образования, существуют формы этих структур, которые не обладают подвижностью и играют в клетках особую роль. Первичные реснички представляют собой неподвижные органеллы, обнаруженные практически во всех клетках позвоночных, исключая клетки крови. В отличие от подвижных ресничек, у клеток обычно находится только одна неподвижная ресничка. На рисунке ниже представлен наиболее типичный пример. Аксонема первичной реснички не содержит центральных микротрубочек, и поэтому ее структура часто обозначается как «9+0». Снаружи большинство первичных ресничек выглядят как обычные реснички. Они начинаются на поверхности клеток и имеют вид простого короткого волоска. Однако, у некоторых высокодифференцированных типов клеток дистальный участок первичной реснички сильно вытянут и превратился в специальный домен, размеры которого сравнимы с размерами самой клетки. Это, например, имеет место в клетках палочек и колбочек, которые содержат фоторецепторы, участвующие в восприятии света сетчаткой глаза. В палочках кончики ресничек вытянуты и образуют большой домен, называемый наружным сегментом, который содержит стопки мембранных дисков, заполненные белковым фоторецептором, родопсином. На рисунке ниже представлен соответствующий пример. Основание первичной реснички соединяет наружный сегмент с остальной частью клетки, аксонема лишь немного вдается в область, где начинается наружный сегмент. За счет транспорта IFT типа, мембранные везикулы, содержащие родопсин, перемещаются из тела клетки в наружный сегмент, и вероятно, это необходимо для его образования и функционирования.
Использование наружного сегмента палочки в качестве светового сенсора может иллюстрировать широкое распространение такой возможности среди первичных ресничек. Возможно, что подобная функция первичных ресничек только начинает исследоваться. В других типах клеток содержатся более примитивные первичные реснички, в которых могут быть локализованы различные типы рецепторов. Локализация рецепторов в первичных ресничках может превратить их в некое подобие антенн, которые могут улавливать изменения в окружении и передавать эту информацию в клетку.
Некоторые редкие болезни человека связаны с мутациями , в результате которых реснички и жгутики утрачивают подвижность. Больные, которым эти мутации передаются по наследству, обычно страдают хроническими респираторными инфекциями, поскольку неподвижные жгутики не способны выводить из дыхательных путей слизь, а также захваченные патогенные микроорганизмы и раздражители. Больные мужчины часто оказываются бесплодными, так как сперматозоиды у них лишены подвижности. К числу широко известных заболеваний, связанных с неподвижностью ресничек и жгутиков, относится синдром Картагенера. Наряду с респираторными инфекциями и бесплодием, у половины больных отмечается извращенное расположение внутренних органов, при котором изменяется нормальная левосторонняя их асимметрия. Предполагается, что на одном из этапов раннего развития, до момента закладки внутренних органов, в результате биения жгутиков, в эмбрионе создается циркуляция жидкой среды, и возникает градиент секретирующих-ся веществ, способствующих морфогенезу, что определяет левостороннюю асимметрию. В отсутствие морфогенного градиента органы занимают случайное положение слева и справа. У мышей с мутациями по флагеллярным динеинам или моторам ответственным за IFT, также отмечается извращенное расположение внутренних органов. Это говорит о том, что мутации, влияющие на подвижность жгутиков или на их образование, могут приводить к порокам развития.
Выразительность
Точность
Полнота
Критерий, используемый для оценки отчета об исследовании; конкретно в отношении того, предоставляется ли в отчет вся информация, в которой нуждаются читатели, на понятном им языке
Критерий, используемый для оценки отчета об исследовании; конкретно в отношении того, логична и информационно точна ли аргументация отчета.
(отчет может быть неточным: малоответственная обработка данных, нелогичность аргументации , неумелое построение фраз).
Ясность
Критерий, используемый для оценки отчета; конкретно в отношении того, точна ли фразеология отчета.
Применять простое правило:
Отчет должен быть строго организован:
- определить цель отчета;
- общий набросок главных акцентов;
- расставить акценты в логическом порядке;
- определить предмет исследования.
(убрать жаргон, использовать короткие слова, писать просто и естественно, избегать нечетких определений)
Критерий, используемый для оценки того, насколько живо и непосредственно написан отчет
Структура отчета
- титульный лист
- содержание
- краткий обзор
- введение
- основная часть
- заключительные рекомендации
- приложение
Большинство бактерий передвигаются при помощи жгутиков . Рассмотреть жгутики можно только в электронном микроскопе . В световом микроскопе без специальных методов обработки отдельные жгутики не видны.
По расположению и числу жгутиков на поверхности клетки бактерии подразделяются:
На монотрихи - имеют один жгутик (например, бактерии родов Caulobacter и Vibrio );
. лофотрихи - имеют на одном или на обоих полюсах клетки пучок жгутиков (например, бактерии родов Pseudomonas , Chromatium );
. амфитрихи - имеют по жгутику на обоих полюсах клетки (например, бактерии рода Spirillum );
. перитрихи - большое количество жгутиков, располагающихся по всей поверхности клетки (например, бактерии вида E.coli и рода Erwinia ) (рис. 1).
Жгутики представляют собой спирально закрученные нити, состоящие из специфического белка флагеллина . Флагеллин построен из субъединиц с относительно малой молекулярной массой. Субъединицы располагаются по спирали вокруг внутреннего свободного пространства. Аминокислотный состав флагеллина у разных видов бактерий может варьировать.
Монотрих Лофотрих
Амфитрих Перитрих
Рисунок 1 - Типы жгутикования у бактерий
Жгутик состоит из трех частей: нити, крюка и базального тельца (рис. 2). С помощью базального тельца, в которое входит центральный стержень и кольца, жгутик закреплен в цитоплазматической мембране и клеточной стенке. Количество колец у грамотрицательных и грамположительных бактерий различно. У грамотрицательных бактерий имеются четыре кольца: L, P, S, M. Из них L и P - наружная пара колец; S и M внутренняя пара колец. L-кольцо закреплено в наружной мембране, P - в пептидогликановом слое клеточной стенки, S - в периплазматическом пространстве, а M - в цитоплазматической мембране. У грамположительных бактерий базальное тельце устроено проще. Оно состоит только из двух колец: S и M, т. е. только из внутренней пары колец, которые размещаются в цитоплазматической мембране и клеточной стенке.
Жгутики бактерий по характеру работы подобны корабельному винту. Если клетка имеет много жгутиков, они при движении собираются в пучок, который образует своеобразный пропеллер. Пучок жгутиков, быстро вращаясь против часовой стрелки, создает силу, заставляющую бактерию двигаться почти по прямой линии. После того как направление вращения жгутиков изменяется, пучок расплетается и клетка останавливается, вместо поступательного движения она начинает хаотически вращаться, ее ориентация изменяется. В тот момент, когда все жгутики бактерии снова начнут синхронно вращаться против часовой стрелки, образовав пропеллер, толкающий бактерию, направление ее поступательного движения будет отличаться от первоначального. Таким способом бактерия может изменять направление своего движения.
Рисунок 2 - Структура жгутика
Так как у грамположительных бактерий наружная пара колец отсутствует, то считают, что для вращения жгутиков необходимо наличие только внутренней пары (кольца S и M). Эти кольца, соединенные с вращающимся стержнем, выступающим наружу, и образуют так называемый электромотор, обеспечивающий движение жгутика (рис. 2). На периферии кольца М находятся белки MotB. Белки MotА встроены в цитоплазматическую мембрану и примыкают к краям колец M и S. Вращающий момент возникает за счет взаимодействия субъединиц белка MotВ с белковыми субъединицами MotА. В белковых субъединицах MotА имеется по два протонных полуканала. Через эти протонные полуканалы переносятся протоны из периплазматического пространства в цитоплазму бактерий (подобно протонному каналу АТФ-синтазы). В результате переноса протонов через белки MotА и MotВ происходит вращение кольца М. Установлено, что один полный оборот кольца М связан с переносом через мембрану около 1000 протонов. Таким образом, в качестве источника энергии для вращения жгутиков используется протондвижущая сила, возникающая в цитоплазматической мембране.
Для подвижных бактерий характерны таксисы , т. е. направленная двигательная реакция в ответ на определенный фактор. В зависимости от природы различают хемотаксис , фототаксис, магнитотаксис и вискозитаксис.
Хемотаксис - движение бактерий относительно источника химического вещества. Для каждого микроорганизма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы, или эффекторы. Среди эффекторов выделяют: аттрактанты - вещества, которые притягивают бактерии; репелленты - вещества, которые отпугивают бактерии.
Фототаксис - движение к источнику света или от него, свойственное фототрофным бактериям.
Магнитотаксис - способность бактерий передвигаться по силовым линиям магнитного поля Земли или магнита. Выявлен в клетках бактерий, содержащих магнитосомы и распространенных в водных экосистемах разного типа.
У ряда бактерий выявлен вискозитаксис - способность реагировать на изменение вязкости раствора и передвигаться в направлении ее увеличения или уменьшения. За чувствительность бактерий к градиенту концентраций определенных факторов ответственны специфические рецепторы. Рецептор реагирует на эффектор и передает сигнал определенного типа на базальное тельце жгутика.
Ворсинки (или фимбрии)
Ворсинки, или фимбрии , - поверхностные структуры, которые состоят из белка пилина и не выполняют функцию движения. По размерам они короче и тоньше жгутиков. Число фимбрий на поверхности клетки колеблется от 1-2 до нескольких тысяч, их имеют как кокковидные, так и палочковидные бактерии. Различают два типа фимбрий: общие и специфические.
Фимбрии общего типа выполняют функцию прикрепления клетки к поверхности субстрата. Не исключается возможность их участия в поступлении крупномолекулярных соединений в цитоплазму клетки.
Специфические ворсинки - половые пили, обнаруженные у клеток так называемых доноров, т. е. у клеток, содержащих половой фактор (F-плазмиду) или другие донорспецифические плазмиды. Если в клетке бактерий находится половой фактор, то на их поверхности синтезируются одна-две половые F-пили на клетку. Они имеют вид полых белковых трубочек длиной от 0,5 до 10 мкм. F-пили играют определяющую роль в образовании конъюгационных пар при переносе генетического материала от клетки донора в клетку реципиента.